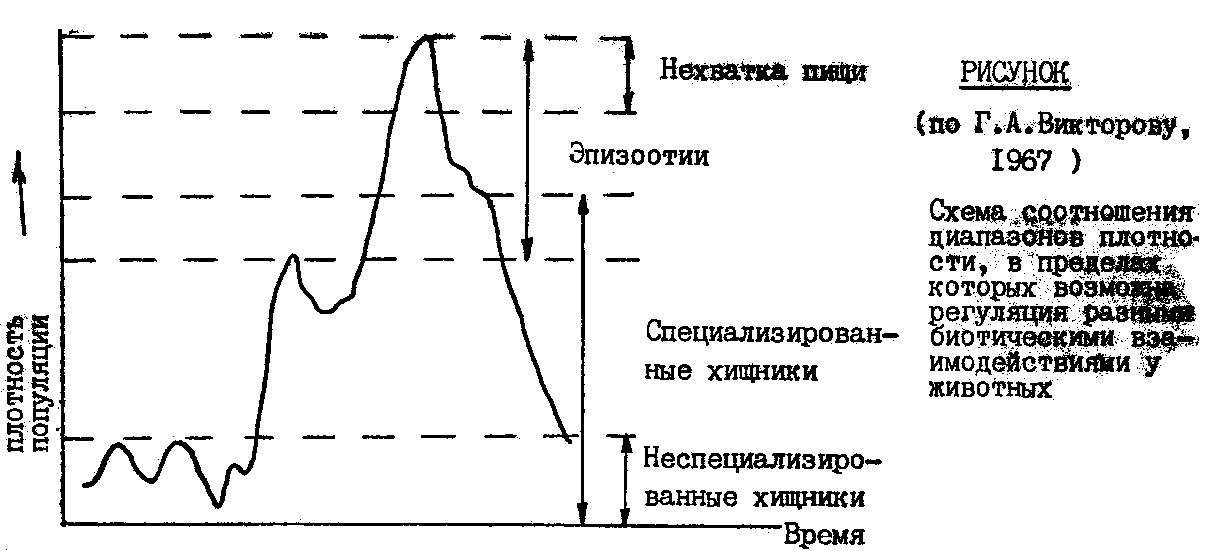
Схема соотношения диапазонов плотности, в пределах которых возможна регуляция разными биотическими взаимодействиями у животных (по Г.А.Викторову, 1967 )
Ответы на вопросы находятся в конце файла
1. Как Вы думаете, с помощью каких механизмов может осуществляться регуляция плотности популяции у животных?
2. Предложите как можно больше способов, которые могли бы использовать бактерии для защиты от вирусов.
3. Во всемирно известном Институте Желтка (г. Дедмихейск-на-Оке) из эукариотических клеток была выделена обратная транскриптаза (белок, который синтезирует ДНК, используя в качестве матрицы и-РНК). Как бы Вы объяснили наличие обратной транскриптазы у эукариот?
4. У растения семейства крестоцветных обнаружены гены А, В и С,
мутации которых ведут к:
А: появление чашелистиков вместо лепестков венчика;
В: появление лепестков вместо тычинок;
С: появление тычинок вместо пестиков.
Как Вы думаете, какова функция этих генов в норме? К чему может
привести наличие в геноме одной особи двух или трех мутантных аллелей
этих генов?
5. Почему одноклеточность накладывает ограничения на размеры организма? Если можете, приведите примеры очень крупных одноклеточных и объясните, как им удается ослабить эти ограничения.
6. Известно, что животные могут менять цвет всего тела или отдельных его частей. Предложите как можно больше механизмов изменения окраски. Если можете, проиллюстрируйте каждый из предложенных механизмов примерами.
7. Как определить, в какой из хромосом находится интересующий нас ген?
1. У большинства животных самки живут дольше самцов. В чем может заключаться биологический смысл этого явления? Возможна ли обратная ситуация? Если да, то в каких случаях? Если нет, то почему?
2. При попытке применения биологических методов борьбы с капустной белянкой (выпуск на поля наездника, личинки которого паразитируют в гусеницах этой бабочки, что приводит к гибели гусениц) повреждения капусты оказались ещё большими. С чем это может быт ь связано?
3. Каковы, по-Вашему, функции пигментов /красящих веществ/ животных и растений? Ответ обоснуйте, если можете, приведите примеры.
4. Палеонтологам часто приходится восстанавливать облик вымершего животного и некоторые особенности его образа жизни по разрозненным остаткам отдельных частей скелета. Какими принципами руководствовались бы Вы, если бы Вам пришлось решать такую задачу? Какие ошибки и почему могут здесь возникнуть?
5. Почему разные млекопитающие имеют разную длительность беременности? Какие факторы и каким образом могут влиять на длительность беременности?
6. Известно, что в тропиках среди общего числа видов высших растений высока доля вечнозеленых. В средних широтах она резко снижается, а в арктических возрастает, но не так сильно, как в тропиках. Предложите и аргументируйте свои объяснения этого факта.
7. Известны случаи, когда съедобные виды насекомых очень похожи по внешнему виду на несъедобные (ядовитые, жалящие, дурно пахнущие и пр.), что уменьшает вероятность поедания хищниками. Но известны также случаи, когда несколько видов насекомых очень похожи друг на друга, и не удается обнаружить похожего на них несъедобного вида. Как Вы думаете, чем можно объяснить такую ситуацию?
1. У многих копытных самцы и самки за исключением периода спаривания используют различные местообитания. В чем может заключаться биологический смысл этого явления? Предложите и обоснуйте как можно больше гипотез.
2. Как животные могут защищаться от своих паразитов?
3. С какими особенностями образа жизни связано наличие у различных животных приспособлений типа крючков и присосок?
4. По сообщению инопланетного разума, записанному контактёром, многочисленные отравления съедобными грибами летом 1992 года были вызваны тем, что они заражены так называемыми грам-положительными бактериями, которые при попадании в желудок человека начинают выделять инсулин. Лечение - введение внутривенно сыворотки против грам-положительных бактерий. Как Вы считаете, что здесь разумно, а что - нет? Почему?
5. У разных животных с двусторонней симметрией одноименные органы, расположенные справа и слева, часто отличаются по форме и(или) размеру. С чем это может быть связано?
6. "Головокружение случается от разных причин, как то: от голода, при резком подъеме или спуске, в горах, от кружения на карусели, от потери крови, у беременных и, наконец, от успехов. Это хороший пример того, как весьма отличные воздействия запускают один и тот же физиологический механизм", - считает физиолог доктор Литвинус. Каковы, по-Вашему, физиологические механизмы этого явления в каждом из случаев? В чем их сходство и различие? Согласны ли Вы с доктором?
7. Однажды знакомый архивариус показал Алексею Михайловичу
Чебуркову старинную рукопись, в которой было написано:
В чистом поле ни кусточка -
Только белый гриб родился.
Две зимы рос и два лета,
Начал спорами кидаться,
Споры эти полетели.
Встретились с водорослями,
С ними выросли в лишайник,
Что забрался на березу,
Прокусил кору коварно
И сосал березы соки.
Тихо плакала береза,
Сок соленый источая
На опята, что под нею
Притулилися на травке,
И на плаун, зацветавший
Скромным цветом у березы.
Вдруг вдали заржали кони ...
На этом рукопись обрывалась.
- Я думаю, это дошедшее до нас начало записи неизвестной народной
былины, - сказал архивариус. - А я думаю, что это фальшивка, притом
изготовленная относительно недавно плохо образованным человеком, -
возразил А.М.Чебурков. Согласны ли Вы с Чебурковым? На каком основании
пришел он к такому выводу?
1. Многие животные и растения могут отделять от себя некоторые органы и части тела. Какое значение для них это может иметь?
2. Как птицы и млекопитающие спасаются от непогоды?
3. Почему у сидячих морских животных встречаются яркие расцветки?
4. С какими особенностями образа жизни может быть связано наличие на одном растении одновременно нескольких типов листьев, разных по форме и строению?
5. Каковы могут быть причины миграций?
6. Какими способами животные добывают пищу, которую не могут увидеть?
7. В одном из романов конца XIX века можно прочитать следующее:
"... На следующий день отставной поручик Алексей Михайлович Чебурков
зашел в гости к соседу, известному коннозаводчику и местному стихотворцу
Козлову, слывшему в округе большим любителем животных. К его
приходу хозяин как раз окончил новое стихотворение, кое и поспешил
прочесть гостю:
Жалко мне мою лошадку,
Когда злой четверокрылый
Овод кровь сосет коняге,
Жалко мне мою собачку,
Что потеет очень сильно
После утренней охоты.
Лучше тихо на рыбалке
Наловить в пруду форелей
И угрей в наряде брачном.
Как красив мой пруд тенистый!
Тихо ползает по веткам
Губка желтая - бадяга,
Объедая с них беззубок.
Иногда, бывает, словит
Водяного скорпиона
С грозным жалом ядовитым,
Но не страшен он бадяге...
Чуть подальше, над тростинкой,
Стрекоза с зеленой мухой
Друг за дружкою летают,
Веселясь в игре кипучей...
Но я что-то засиделся!
Ведь пора мне в лес тенистый,
Чтоб послушать коростеля
Да набрать для попугая
Можжевеловых орешков.
- Да у Вас, батенька, воспаленная фантазия! - воскликнул поручик. -
Нагородить столько небылиц в одном стихотворении!"
Согласны ли Вы с А.М.Чебурковым? Какие конкретные места в стихотворении
имел он в виду?
1. Некоторые животные едят у растений только цветки. С чем это может быть связано?
2. Для чего многие животные сооружают не одно гнездо (нору), а несколько? Приведите примеры.
3. Известно, что покровы многих динозавров были снабжены такими образованиями, как пластины, шипы, рога, гребни. Как Вы думаете, какие функции могли выполнять эти образования?
4. Стебли многих растений после опыления удлиняются и разрастаются. Как Вы думаете, чем это может быть полезно для растения?
5. Какие функции могут выполнять щетинки и волоски у насекомых?
6. Каким образом распашка луга может привести к снижению численности некоторых видов растений на соседнем лугу?
7.
"...Я видел лес,
Который рос из моря.
Там вместо птиц
На ветках сидели рыбы.
Я видел места,
Где обычные травы,
Вырастая,
Скрывают всадника.
Я видел озёра,
В которых слепые
Белые рыбы
Ловили слепых раков.
Я видел птиц,
Уносивших черепах
И бросавших их на скалы
Чтобы разбить панцирь.
Я видел ручьи
С кипящей водой,
Пахнущей серой,
И в них росли водоросли.
Я видел дерево Ко,
Цветущее чёрной ночью
Белыми
Светящимися цветами.
Но нигде не видал я
Конца своего пути."
Где побывал путешественник? Что он видел? Имеются ли в его рассказе
неточности либо вымысел?
Вопрос 1. Растения чаще всего отделяют свои части /органы/ в связи с размножением. Плоды семена и споры - наиболее очевидный пример. Многие растения имеют ломкие побеги, что способствует вегетативному размножению, а иногда - расселению. Например, "перекати-поле" /это не вид, а жизненная форма, характерная для многих степных видов/. Клубки обломанных побегов этих растений переносятся ветром на большие расстояния, рассеивая по дороге семена. Иногда на взрослом растении формируются и потом обламываются молодые проростки (напр., каланхоэ). В тропических лесах, где прорастание из семян часто затруднено, проростки еще на дереве достигают крупных размеров и отращивают крепкий стержневой корень, после чего отрываются, втыкаются в почву и укореняются.
Второе распространенное явление - листопад. Кроме листьев, растения иногда сбрасывают ветви (напр. у ряда хвойных и у саксаула отпадают укороченные однолетние побеги). Роль листопада - уменьшение испарения воды во время сухого сезона, а также сброс ненужных органов на период покоя, а вместе с ними и накопленных вредных продуктов обмена (функция выделения). Видимо, с той же целью некоторые растения /земляничник, пузыреплодник/ каждый год сбрасывают кору, в которой в спец. клетках и полостях накапливаются метаболиты. Отбрасываются также уже отслужившие свое и ставшие ненужными частицы /кожура семени с проростка, лепестки цветка/. В сверхурожайные годы растения могут терять часть завязей плодов, на нормальное развитие которых им не хватило бы "сил".
В крайних условиях существования /тундра, горные склоны/ встречается явление партикуляции: кустарники и деревья при резком ухудшении условий могут распадаться на части. В середине ствола образуется слой перидермы и изолирует чати части друг от друга. Это простой и экономичный способ размножения, после которого выжившие могут размножаться обычными способами.
Причины отделения органов и частей тела у животных также могут быть самыми разными. Разберем лишь некоторые из них. Это и аутотомия (самокалечение), когда в целях защиты от хищника орган или часть тела отделяется (хвост у ящериц, задняя часть кишечника у голотурий и пр.), а затем регенерирует. Это избавление от отслуживших свое и ставших ненужными органов /крылья у половых особей муравьев и термитов, плацента млекопитающих, отторгающаяся при родах/, в том числе редкие случаи катастрофического метаморфоза (отбрасывание эктодермального слоя - пилидия - у личинок немертин или эктодермальной ленты жгутиковых клеток у плутеусов - личинок иглокожих).
Подавляющее большинство случаев утраты личиночных органов не являются ответом на вопрос, ибо происходит не отбрасывание, а постепенная резолюция и исчезновение органа (хвост у головастика и т.п.). Не годятся также различные случаи линьки, выстилания гнезд пухом или шерстью, продукция половых клеток и т.п., так как все это не соответствует определению "орган или часть тела".
Возможные спорные случаи должны, вероятно, трактоваться в пользу школьника. При аналогах вегетативного раэмножения у животных дочерние особи развиваются иногда из отдельных участков тела материнских особей, предварительно отделившихся /напр., у планарий/. У ленточных плоских червей при размножении отделяются членики (проглоттиды), а у ряда многощетинковых (полихет) существует явление эпитокии, когда взрослый червь "отращивает" на заднем конце тела аналогичную особь с развитой половой системой, которая затем отделяется и участвует в половом размножении, восстанавливая уже после отделения головной конец. Бывают и вовсе необычные случаи, напр. у самцов головоногих моллюсков одно из щупалец разрастается и образует так называемый гектокотиль, который наполняется половыми продуктами. У некоторых видов гектокотиль может отрываться и самостоятельно плавать в поисках самки. Встретив самку, он проникает в ее мантийную полость и производит оплодотворений.
ВОПРОС 2. Во-первых, от непогоды позволяет спасаться устройство покровов тела. У млекоптающих шерсть растет от спины к животу, что способствует отеканию воды и сваливанию снега с тела животного. У птиц перья налегают друг на друга подобно черепице, благодаря чему намокают только кончики перьев, а само тело птицы остается сухим. От намокания спасает также жировая смазка оперения и шерсти. Если же покровы все-таки намокают снаружи, то они слипаются так, что образуется почти не пропускающий воду слой, защищающий подшерсток (пух) и кожу. Распушение шерсти и перьев создает воздушную прослойку вокруг тела животного, препятствующую теплообмену. При ветре и холоде звери и птицы, как правило, принимают позы, минимизирующие поверхность тела и уменьшающие теплоотдачу и увеличивающие обтекаемость (звери сворачиваются в клубок и прижимают уши и хвост, а птицы прижимают голову к груди, плотно складывают крылья и перья хвоста).
Во-вторых, будучи теплокровными, звери и птицы используют для согревания дрожательный термогенез, а для охлаждения - усиленное потоотделение и учащенное дыхание, а также испарение с постоянно влажных поверхностей (напр., полость рта). Подкожный жир используется для теплоизоляции, а запасы бурого жира могут разлагаться с высвобождением тепла для кратковременного разогревания. Наконец, во внешних охлаждающихся частях тела может замедляться кровобращение по сравнению с внутренними.
В-третьих, звери и птицы могут скрываться от непогоды в различных естественных убежищах или создавать свои.
В-четвертых, они могут откочевывать в места, где погода лучше. Морские млекопитающие при кратковременных штормах погружаются в слои, где волнение меньше, а грифы могут скрываться от солнца в тени облаков.
В-пятых, животные, живущие в "скоплениях" /стадах, стаях, колониях/, при непогоде могут сбиваться, уменьшая совокупную теплоотдачу и помогая друг другу иными способами (в особенности, молодняку).
В-шестых, млекопитающие часто способны впадать в спячку либо просто уменьшать активность, сокращая расход энергии. Самые мелкие птицы - колибри - при понижении температуры среды также впадают в кратковременное оцепенение.
ВОПРОС 3. Многие губки, кишечнополостные, асцидии и некоторые моллюски обладают яркой окраской, хотя, ввиду сильного поглощения толщей воды определенных /в первую очередь красной/ частей спектра и общего резкого падения освещенности с увеличением глубины, лишь в мелководных биотопах они действительно выглядят яркими, а на глубине уже в 20 м красная, к примеру, актиния выглядит зеленовато-серой. В некоторых случаях яркая окраска является предупреждающей, но встречается широко и у интенсивно поедаемых и, наоборот, явно несъедобных видов, а также у таких, хищники которых слепы. В мелководных биотопах яркая окраска может быть маскирующей /напр., среди красных и зеленых водорослей/; то же касается преобразованной яркой окраски на глубине /серо-зеленая или голубоватая среди донных включений на общем темно-сером фоне дна/. В ряде случаев сами животные могут быть не окрашены, часто полупрозрачны/, а цвет им может придавать проглоченная пища (вариант: пигменты из жертв накапливаются и откладываются в покровах), симбионты /зоохлореллы и зооксантеллы, напр., у кораллов/, дыхательные пигменты, которые имеют яркий цвет из-за наличия металлсодержащего активного центра /медьсодержащие голубые гемоцианины, железосодержащие зеленые хлоркруорины и красные гемоглобины и т.п./, окрашивают в яркий цвет кровь /гемолимфу/ животных, которая и просвечивает сквозь покровы. Многие губки и моллюски обрастают снаружи ярко окрашенными, часто корковыми водорослями, которые используют их просто как удачный субстрат. В некоторых случаях яркая окраска может быть атавизмом, сохранившимся с тех времен, когда она была функциональной. В целом формулировка вопроса допускает как ответы, в которых сама окраска несет функциональную нагрузку, так и ответы, в которых окраска есть производное от строения и образа жизни организма.
ВОПРОС 4. Разнообразие форм листьев на одном и том же растении
носит название гетерофиллии (разнолистности). Примеры такого разнообразия:
1) листовая мозаика - разные форма и размер листьев связаны с
приспособлением к более эффективному поглощению света.
2) возникновение разнолистности в разных условиях: при обилии света
у молодого побега плюща развиваются нормальные пятиугольные листья с широкой
листовой пластинкой, в то время как у затенённого побега листья мелкие и
овальной формы. У филлодийных акаций лист представлен уплощенным зелёным
черешком, на котором сидит двойная сложнорасчленённая листовая пластинка.
В условиях засухи эта пластинка отпадает, и в дальнейшем растение имеет на
себе лишь черешок листа, тем самым уменьшая потерю влаги в организме.
3) Листья могут выполнять различные нетипичные функции:
а. прицветные верхушечные листья привлекают насекомых-опылителей к
соцветию /фиолетово-синие листья у иван-да-марьи/
б. у папоротника страусовое перо листья бывают двух типов - одни зеленые,
фотосинтезирующие, другие - коричневые, содержащие спорангии
со спорами /сорусы/
в. защитные образования листового происхождения - колючки у барбариса,
почечные чешуи, корневищные листья у ландыша
г. ловчие аппараты насекомоядных растений
д. у лазящих лиан некоторые листья превращены в усики
4) Разнолистность очень распространена у растений, живущих в двух средах,
например у водных /стрелолист, водяной лютик и т.д./. Подводные листья.
таких растений длинные и узкие, а надводные - с широкой, уплощенной листовой
пластинкой.
ВОПРОС 5. Миграции вызываются различными причинами, простое перечисление примеров еще не является ответом на этот вопрос. Здесь мы рассмотрим основные типы миграций.
Наиболее известны сезонные миграции животных, либо уходящих от неблагоприятных условий (перелеты птиц, миграции оленей карибу), либо собирающихся с мест откорма в места размножения (ход рыбы на нерест, миграции красных сухопутных крабов в море на многих тропических островах, миграции серых китов из Берингова моря к "родильным домам" у побережья Центральной Америки и т.п.). Последний тип миграций порождает и обратную волну - миграции от мест рождения к местам откорма /скат мальков из рек в море/.
Отдельно можно выделить пищевые миграции. Например, стада травоядных способны очень быстро истощить пастбище, поэтому они все время перемещаются на новые места откорма. Тунцы и киты способны пересекать океан, чтобы добраться до мест, где на границе теплых и холодных течений возникает высокая концентрация корма. Совсем особым типом нерегулярных миграций является особая реакция популяций на перенаселенность, когда огромные массы "избыточных" животных мигрируют с неспособных обеспечить их существование мест /классические примеры: лемминги и перелетная саранча, у которой возникают даже четкие морфологические отличия от оседлых представителей вида/.
Все вышеперечисленные миграции, характерные и для наземных, и для водных животных, относятся к горизонтальным. Для водных животных характерны еще вертикальные миграции, когда они перемещаются вверх-вниз в толще воды. Можно выделить два основных типа вертикальных миграций - суточные (связанные в целом с неодинаковостью абиотических /температура, освещенность, содержание кислорода и пр./ и производных от них биотических характеристик разных слоев воды) и онтогенетические (связанные с наличием расселительной личинки, обитающей в иных, нежели взрослые слоях, или с размножением /откладкой икры/ в иных слоях). При суточных миграциях животные по ночам, как правило, поднимаются вверх, в богатые пищей слои воды, а днем опускаются вниз, спасаясь, например, от хищников /подавляющее большинство планктонных животных и многие рыбы/.
Интересны миграции растений - планктонных водорослей, которые выбирают в зависимости от времени суток подходящие по освещенности и содержанию минеральных веществ слои.
Сезонные вертикальные миграции у наземных животных, живущих в горах, также связаны с неодинаковостью условий на разных высотах над ур. м., но по своим причинам сходны с горизонтальными.
ВОПРОС 6. Во-первых, во многих случаях при добывании пищи используется не зрение, а иные сенсорные системы, как то обоняние /хеморецепция/, осязание /тактильная рецепция/, слух /включая специальные системы локации/, восприятие колебаний /сейсмослух у змей, органы боковой линии у рыб и т.п./, температурная локация /многие змеи, ряд кровососущих насекомых/ и т.д. Интересно обнаружение добычи при помощи искривления собственного электромагнитного поля /электрический угорь/ или по следам жизнедеятельности /дятел по следам личинок/. Хеморецепция используется как животными, в принципе способными видеть добычу /млекопитающие, змеи, охотящиеся в скважинах почвы/, так и животными, не имеющими такой возможности /движение одноклеточных к скоплениям бактерий по градиенту концентраций определенных веществ/.
Во-вторых, часто используются пищевые частицы весьма малого размера и/или диспергированные в среде, и зрение либо отсутствует вовсе, либо используется для других целей. Так бывает у паразитов, всасывающих вещества всей поверхностью тела или поедающих то, что находится вблизи места прикрепления. Сюда же относятся различные фильтраторы /от инфузории до кита/, детритофаги поедающие органику грунта, соскабливатели /напр., брюхоногие моллюски/. Они могут быть более или менее избирательными в связи со строением питательного аппарата и наличием или отсутствием предварительного поиска скоплений пищи. Так, инфузория-туфелька неизбирательно поглощает все частицы определенного размера /размер определяется размером "рта"/, а переваривает лишь часть органических частиц. Сходным образом поступают неизбирательные детритофаги типа полихеты пескожила, поглощающие грунт, пропускающие его через кишечник и переваривающие органику, а остальное выбрасывающие. Другие детритофаги /некоторые моллюски/ предварительно производят поиск скоплений пищи, а многие фильтраторы кроме того имеют сложно устроенный аппарат, дифференцирующий захваченные частицы по размеру, плотности, вкусу и т.п. и отбрасывающий ненужное. Многие животные постоянно поедают часть своих внутриклеточных /инфузории, гидры - одноклеточных водорослей/ или полостных симбионтов /в кишечнике термитов переваривается часть симбиотических жгутиконосцев гипермастигот, и это нормальный процесс, поскольку таким образом термиты усваивают растительную клетчатку, которую сами по себе усваивать не могут, а кормят ею симбионтов, поедая затем часть наращиваемой ими биомассы/. Существуют также "ловушечники" типа муравьиного льва, личинки которого поедают всех попавших в ловушку не слишком крупных насекомых, не видя их.
ВОПРОС 7. Основные несообразности в тексте:
1. Оводы - насекомые отряда двукрылых и, соответственно, имеют одну пару
крыльев (вторая, рудиментарная, пара превращена в жужжальца).
2. Оводы не сосут кровь и вообще лишены ротового аппарата. Их самки
откладывают яйца или, чаще, отрождают живых личинок во внутренние полости
(ушные и носовые проходы) или на кожу животных, на которых оводы
паразитируют.
3. У собак практически нет потовых желез и потому они не могут сильно
потеть, испарение происходит у них гл. обр. через полость рта.
4. В природе форели, как и большинство лососей, живут в быстро текущих
водах или крупных олиготрофных озерах, а никак не в прудах. Вместе с тем при
промышленном разведении мальков форели иногда подращивают в прудах при
соблюдении комплекса зоотехнических мероприятий, но в имении коннозаводчика
Козлова в конце XIX века это маловероятно.
5. Зато "угрей в наряде брачном" в пруду встретить невозможно. Брачный
наряд у угрей, конечно, есть, но "одевают" они его на заключительной стадии
миграции - в Саргассовом море, где размножаются. Личинки европейского угря
мигрируют с Гольфстримом к берегам Европы, где молодые угри входят в реки,
достигая там размеров взрослых особей. Неполовозрелый угорь может попасть
и в расположенный в верховьях реки пруд, проникнув туда по водотоку или по
неширокой влажной плотине (т.е. по суше).
6. Мелкая придирка - имеются в виду, естественно, затонувшие в пруду
ветки.
7. Бадяга, как и все взрослые губки, неподвижна /прикреплена к субстрату,
напр., к той же ветке/ и питается, отфильтровывая мелкие органические частицы,
а никак не крупных двустворчатых моллюсков - беззубок. Кроме того, бадяга
буро-зеленого или бурого цвета, а не желтого. Беззубки живут на дне водоемов,
наполовину зарывшись в грунт. Пруд, чтобы в нем жили беззубки, должен быть
довольно крупных размеров, и желательно проточный, т.к беззубки требовательны
к содержанию кислорода, не выносят слишком толстого слоя ила и для
размножения нуждаются в рыбах, на которых развиваются и которыми
переносятся их личинки - глохидии.
8. Водяной скорпион - представитель сем. непид отряда клопов. Он не имеет
вытянутого брюшка и жала и никакого отношения к настоящим скорпионам. На конце
брюшка у него вытянутая воздухоносная трубочка, которую он периодически
выставляет над поверхностью воды и через нее дышит, он питается некрупными
движущимися объектами (рачками, личинками насекомых, мальками),
и действительно не страшен бадяги.
9. Стрекозы, как известно, питаются мухами, поэтому коннозаводчик Козлов
мог наблюдать именно охоту, а не игру.
10. Коростель не живет в лесу - он живет на заболоченных лугах.
Издаваемые самцами коростеля в брачный период звуки трудно назвать
приятными для слуха, они похожи на скрежет /дёргание"/, хотя о вкусах не
спорят. Поскольку коростели "поют" в предрассветные часы, их невозможно
услышать "засидевшись" у пруда и пронаблюдав то, о чем рассказано в
стихотворении выше, если только не "засидеться" у пруда до самой ночи...
11. Можжевельник, будучи голосеменным растением, имеет не орешки
/так называются плоды некоторых цветковых растений/, а шишки с утолщенными
мясистыми чешуями, которые иногда не вполне правильно называют "шишкоягодами".
Это женские шишки можжевельника, содержащие каждая по три семени. Они имеют
сладковатый вкус и в природе поедаются многими птицами, например, дроздами,
вполне охотно. Возможно, их ел и попугай коннозаводчика Козлова, хотя в
природе попугаи и можжевельники, насколько известно, вместе не встречаются.
ВОПРОС 1. Использование различных местообитаний (М) доказано для
самок и самцов благородных, американских и северных оленей, серн,
толсторогих баранов и некоторых других копытных. Адаптивные преимущества
такого поведения обсуждаются в настоящее время в рамках, по меньшей мере,
пяти гипотез:
1) альтруизм самцов, оставляющих самкам и молодняку лучшие
/по разным показателям, возможны варианты/ М;
2) избегание хищников самцами, истощенными во время гона;
3) сведение к минимуму агрессивности между самцами из-за самок
в период покоя;
4) выбор самцами открытых М чтобы избежать повреждений рогов в период
их роста;
5) оптимизация использования кормов самцами и самками с молодняком.
Школьниками могут быть предложены и иные разумные гипотезы, либо совсем новые, либо возникшие как результат развития или частичного совмещения вышеперечисленных. В целом, критерии различной избирательности самок и самцов - следствие их разных стратегий размножения. Избирательность М у самок может определяться потребностями в воде в период кормления молодняка, концентрацией и качеством кормов, а также защитными условиями для избегания хищников. Существенными могут оказаться для самок и требования молодняка к качеству стартовых растительных кормов, удаленности солонцов и т.п. По мере роста молодняка эти требования, а также поведение самок и характер использования М могут меняться. У самцов репродуктивный успех во многом определяется физическим состоянием и весом тела. Поэтому зимой, например, они более нуждаются в усваиваемой энергии, чем в белке, и сильнее используют иные источники корма, чем самки, у которых высоки именно потребности в белке в связи с беременностью. Оптимальное кормодобывание самцов предполагает, что они должны преимущественно использовать высококачественные, но временные корма и избегать обедненных кормами М, что с неизбежностью ведет к увеличению двигательной активности в поисках подходящих М и меньшей защищенности от хищников по сравнению с самками, но и одновременно компенсирует энергозатраты.
В целом школьникам предлагается указать как можно больше межполовых различий у копытных /морфологических, физиологических, поведенческих и пр./, связанных непосредственно с биологией размножения, и далее объяснить, как с учетом этих различий разные полы могут оптимизировать свои отношения со средой обитания, подобрав условия этой среды.
ВОПРОС 2. Животные защищаются от паразитов разнообразными способами, которые условно можно разделить на две группы: 1) "активная" защита - гл.обр. поведенческие реакции и 2) "пассивная" защита - особенности строения, физиологии и биохимии организма.
1) "Активная" защита:
а. удаление наружных паразитов:
- самостоятельно с помощью грязевых и песчаных ванн, трением о стволы деревьев, механически с помощью конечностей, зубов, хвоста и т.п.
- симбиотические и внутривидовые связи /рыбы-"чистильщики", выискивание/
- заход в зоны, неблагоприятные дяя жизни паразитов /киты заходят на мелководья с опресненной водой, где погибают и отваливаются их океанические наружные паразиты/
- использование межвидовых связей /млекопитающие и птицы, ложащиеся в муравейник, очистка птицами от паразитов крупных копытных и т.п./
б. поедание вредных для внутренних паразитов веществ /напр., лекарственных растений/
в. активность в периоды, когда паразиты отсутствуют
г. дезинфекция и дезинсекция убежищ /напр., птицами - своих гнезд с помощью растений, химических веществ (извести), выбрасывания из гнезда помета и старой подстилки с живущими там личинками паразитов и т.п./
2) "Пассивная" защита:
а. особые строение покровов /толстая кожа, линька, при которой
сбрасывается часть паразитов и т.п./
б. выделение ядовитых веществ кожей /некоторые рыбы, амфибии/, наличие
токсичных для ряда паразитов продуктов обмена
в. функционирование иммунной системы /фагоцитоз, продукция антител,
действие системы компплемента крови и пр./
г. частичная защита в результате нормального функционирования систем
организма /гибель части паразитов от ферментов желудочно-кишечного тракта,
особенно тех из них, которые далее должны проникнуть в более специфичные
для них места паразитирования; выведение из организма с продуктами обмена
части паразитов пищеварительной и мочеполовой систем и т.п./.
Сюда же можно отнести поддержание целостности и нормального функционирования
органов, кожных покровов и пр. как своего рода профилактика /некоторые внешние
паразиты внедряются только на поврежденные участки покровов; другие, напр.,
энтамёбы, являются в норме комменсалами и вызывают амебиоэ - изъязвления
стенок кишечника - лишь в некоторых случаях, в том числе при возникновении
язв иного происхождения, нарушениях нормальной перистальтики и других
патологиях/
д. изоляция паразитов, особенно тканевых, специальными уплотнениями,
сужающая "поле деятельности" паразита и уменьшающая воздействие на организм
хозяина токсичных продуктов его обмена и распада ткани /напр., изоляция
личинок цестод в мышцах промежуточных хозяев разрастанием соединительной
ткани и т.п./
е. при поражении вирусами клетки высших животных вырабатывают белок
интерферон, подавляющий синтез и-РНК, а поскольку подавляющее число
синтезирующихся в зараженной клетке и-РНК служат для синтеза вирусных белков,
то вирус не может размножиться и гибнет
ж. избирательная рецепция клетками определенные веществ и избирательная
прницаемость для разных веществ их мембран /особенно в клеточных пластах,
где плотные межклеточные контакты исключают транспорт по межклеточным
пространствам, а внешняя поверхность пласта покрыта гликокаликсом/ создает
преграды для перемещения по организму вредных продуктов деятельности
паразитов, а в сочетании с лизосомальной активностью клеток - и самих мелких
/одноклеточных/ паразитов
Возможны, конечно, и другие, не указанные выше варианты ответов.
ВОПРОС 3. Для удобства можно рассмотреть несколько групп вариантов использования подобных приспособлений:
1. Использование крючков и присосок для удержания на какой-то поверхности. Примеры: паразитические формы /сосальщики имеют на головном конце и на брюшной стороне присоски, с помощью которых присасываются к стенкам кишечника хозяина; многие ленточные черви используют для той же цели крючки на сколексе; использование и крючков, и присосок наблюдается у многоусток - паразитов рыб/, лазающие формы /у большинства мух и квакш - присоски на лапах/, многие комменсалы - для передвижения вместе с хозяином /рыба-прилипала с присоской на верхней стороне головы, актиния с присоской на подошве, но также и с клейким веществом/, животные быстро текущих вод и т.п. Некоторые животные могут использовать определенные органы как присоски лишь иногда /большинство брюхоногих моллюсков - при резких воздействиях используют ногу как присоску большой площади, в то время как движутся либо с помощью ресничек на ней (водные формы), либо скольжением по слою слизи с помощью мускульных сокращений ноги. Для ряда литоральных форм использование ноги как присоски становится правилом, и прижатая к субстрату раковина кроме прочего спасает их от обсыхания в отлив (акмеи, хитоны)/.
2. Крючки часто используются для пробуравливания и продырявливания /шестикрючная личинка свиного солитера своими крючками пробуравливает стенку кишечника и с током крови или лимфы переносится по телу/
3. Для поражения и/или удержания добычи /когти у большинства хищников или аналогичные им крючкообвазные образования, крючкообразные хелицеры паукообразных, присоска на конце языка хамелеона и пр./.
4. Для рытья и рыхления грунта /когти крота и барсука, клыки свиней и пр./
5. Крючкообразные клювы птиц и черепах создают, с одной стороны, наибольшее режущее или давящее усилие, а с другой - способствуют удержанию пищи.
6. Часто крючки или присоски используются для удержания полового партнера /одноустка спайник парадоксальный и многие другие/.
В целом возможно очень много вариантов ответа на этот вопрос, и более высокой оценки заслуживают, видимо, те, в которых делаются некоторые функциональные обобщения, а не просто приводятся конкретные примеры.
ВОПРОС 4. Грам-положительные бактерии - большая группа бактерий, обладающая некоторыми общими свойствами /например, очень сходным строением клеточной стенки/. Все они дают окраску при окрашивании методом Грама - отсюда название. Чтобы ответить на вопрос, всех этих свойств можно и не знать. Хорошо, если кто-то знает, что такие бактерии действительно есть и почему они так называются, но что касается их свойств - неясно, как это можно использовать при ответе. Второе предварительное замечание серьезнее - при хорошем ответе школьник должен понимать, что сыворотка против определенных бактерий - это жидкая часть /плазма/ крови животных или людей, которая содержит антитела /специальные "защитные" белки/ против этих бактерий. После внутривенного введения этой сыворотки зараженным этими бактериями людям в крови происходит встреча этих антител с бактериями и "слипание" /агглютинация/, которая запускает сложную последовательность действий иммунной системы) вызывающую гибель бактерий. Если школьник способен рассказать о работе иммунной системы более подробно и при этом верно - это тоже плюс.
А теперь замечания конкретного характера.
1. Желудочный сок обладает антибактериальными свойствами /не последнюю роль играет здесь низкое значение рН=0,8-1,5/, поэтому неизвестны бактерии, попадающие в желудок и в нем успешно секретирующие токсин.
2. Инсулин в норме секретирует поджелудочная железа млекопитающих, и в природе неизвестны бактерии, могущие делать то же самое. Единственное исключение - бактерия кишечная палочка, в геном которой усилиями генных инженеров введен человеческий ген инсулина, и синтезируемый в специальных лаборатоях большими количествами этих бактерий белок инсулин /который ничем не отличается от обычного/ используется в медицине /напр., лечение диабета/.
Следующие возражения более серьёзны:
3. В желудке выделяются ферменты, расщепляющие белки /напр., пепсин/, а т. к инсулин - белок, то, попав в желудок, может быть расщеплен, утратив свои свойства.
4. Инсулин - не токсин, а естественный гормон человека. Инсулин способствует проникновению глюкозы в клетки печени и депонированию ее там в виде гликогена. Ненормально высокая концентрация его в крови вызывает обычно резкое падение концентрации глюкозы в крови, слабость, обморочное состояние /т.к. в первую очередь в глюкозе нуждается мозг/ и, если не принять срочных мер по повышению уровня глюкозы - смерть. Если бы бактерии действительно могли продуцировать инсулин в желудке, и он, каким-то образом минуя расщепляющие ферменты, проник в кишечник и оттуда всосался в кровь, картина отравления резко отличалась бы от обычного пищевого отравления. Видимо, это скорее всего напоминало бы гипогликемическую кому при введении инсулина диабетикам /при передозировке/.
5. Введение внутривенно сыворотки против грам-положительных бактерий - дело бесполезное. Внутрь желудочно-кишечного тракта она проникнуть не может. Если бактерии могут выделять инсулин только в желудке, а потом проникают в кровь, то там они ничем не опасны. Если же в кровь проникает только инсулин, то сыворотка нужна против него, а не против бактерий - но тут возникает опасность, что эндогенный (вырабатываемый собственно организмом человека) инсулин также вступит во взаимодействие с антителами, инактивиоуется, и возникнет гипергликемическая кома (высокий уровень глюкозы в крови), что тоже без надлежащего вмешательства может привести к смерти.
Замечания 3,4 и 5 составляют ядро правильного ответа на этот вопрос.
ВОПРОС 5. Вообще органы слева и справа формируются обычно в процессе эмбрионального развития одновременно, как правило путем разделения первоначально единой закладки, и точность разделения напополам зависит от случайных причин, такая изменчивость в большинстве случаев не несет функциональной нагрузки и вызвана случайными причинами /она называется флюктуирующей/. Эта изменчивость может усиливаться в процессе дальнейшего развития, поскольку у каждой закладки появляется своя "история" и неодинаковый комплекс воздеиствующих на нее факторов. Кроме того, при ограниченности "ресурса" /ткани начальной закладки/ органы за него конкурируют, и кто получил больше, дальше имеет возрастающее преимущество (автокатализ). Из-за этого реально работают обычно только один яичник и только один семенник. Ассимметрия может быть закреплена генетические как у манящих крабов /одной клешней только кормится и обороняется, другую еще и демонстрирует при внутривидовых контактах/. Здесь уже имеет место быть разделение функций изначально одинаковых органов, которое может быть выгодно, посколько связано со специализацией, позволяющей каждому из органов более эффективно выполнять свою часть функций. В приведенном примере специализация отражается на форме и размере органов, и это различие может быть еще большим вплоть до полной редукции одного из них (напр., одно легкое у змей). Но даже при редукции одного из органов функциональный смысл может быть неясным (так, у птиц редуцирована левая луга аорты, а у млекопитающих правая, это связывают с происхождением этих групп от разных ветвей древних рептилий, но в чем функциональный смысл именно такой редукции - до сих пор непонятно). С другой стороны, разделение функций может не сопровождаться заметной ассимметрией в строении - как у человека с левым и правым полушариями мозга. Ассимметрия в развитии мышц рук у человека, слабо развитая у детей, у взрослых обычно хорошо заметна ввиду преимущественного упражнения одной из рук (то же верно для толчковой ноги у прыгунов и т.п.)
ВОПРОС 6. Разберем каждый из приведенных случаев на доступном школьникам уровне:
1. Голодание приводит к снижению уровня глюкозы в крови, которая является важнейшим энергетическим веществом для клеток мозга - нейронов. Недостаток глюкозы приводит к дисфункции нейронов, сопровождающейся разнообразными симптомами - звоном в ушах, потемнением в глазах, слабостью, головокружением.
2. При резком подъеме или спуске изменение силы тяжести воспринимает вестибулярный аппарат - рецепторный орган, расположенный в полукружных каналах и мешочках внутреннего уха. При разных сильных раздражениях вестибулярного аппарата возникает головокружение, иногда тошнота, рвота.
3. В горах концентрация кислорода в воздухе меньше по сравнению с таковой на уровне моря. Кислород необходим для дыхания клеток, в том числе нейроны очень чувствительны к его недостатку. А дисфункция нейронов.../далее см.1/ Второй механизм головокружения в горах, возможно, связан с синдромом высотобоязни и относится к высшей нервной деятельности человека /функциям мозжечка, коры больших полушарий и лимбической системы/.
4. Кружение на карусели сопровождается инерционным сдвигом эндолимфы, возникающим в полукружных каналах (фрагменте вестибулярного аппарата) при воздействии угловых ускорений. Далее - см. 2.
5. При потере крови уменьшается ее объем в организме и, соответственно, ухудшается кровоснабжение всех органов. Что и говорить, что мозг... /далее см. 1 и 3/
6. У беременных на ранних стадиях развития плода головокружение возникает при токсикозах (причины которых, в общем, заключаются в рассогласовании различных функциональных систем организма в связи с перестройками для жизнеобеспечения плода). Механизм этого явления пока неясен. Идеи школьников на эту тему малопредсказуемы и должны оцениваться исходя из общей здравости и разумности использования знаний по теме "Физиология человека". На более поздних стадиях головокружение иногда может возникать в связи с интенсивным потреблением плодом кислорода и питательных веществ крови и кратковременным голоданием мозга /см. 1 и 3/.
7. Головокружение от успехов проявляется целым рядом неприятных ощущений в виде чувства оглушения, неуверенности при стоянии или сидении, ощущения пошатывания, колебания тела. Это может быть проявлением недостаточности кровообращения в вертебробазиллярной системе, однако чаще носит психогенный характер.
Примечание доктора Литвинуса:"НЕОТЛОЖНАЯ ПОМОЩЬ: Постельный режим. Торекан внутримышечно 1мл или внтрь по 1 драже 3 раза в день. Аминазин в/м по 1 мл 20% р-ра, реглан в/м, седуксен 2 мл 0,5% р-ра. в/м эти же препараты можно назначить внутрь. ПРОФИЛАКТИКА: статья И.В.Сталина "Головокружение от успехов" 3 раза в день до (или после) еды."
ВОПРОС 7. Действительно, эта рукопись составлена весьма безграмотно. Мицелий белого гриба не может развиваться в поле, где нет деревьев. Белый гриб, как и большинство других шляпочных грибов, облигатный микоризообразователь, т.е. не может существовать без симбиоза с высшими растениями и уж тем более образовывать плодовые тела - те самые, которые мы называем грибами и употребляем в пищу. Гифы грибницы оплетают корешки деревьев, частично проникают внутрь, между клетками и в сами клетки, и получают от дерева питательные вещества и гормоны. В свою очередь, гриб помогает корешкам усваивать необходимые вещества из почвы. Белый гриб встречается обычно рядом с елью (светлоголовая форма), сосной (темноголовая форма, боровик), березой (березовая форма со светлой шляпкой и тонкой ножкой), дубом, (серо-бурая шляпка с пятнами, бочонкообразная ножка). Его мицелий может одновременно образовывать микоризу на двух породах деревьев. Плодовые тела обычно образуются каждый год, быстро растут, вызревают и отмирают в течение нескольких дней. В ветренную погоду споры гриба могут пролетать значительные расстояния, но образовать лишайник белый гриб не способен. Лишайник - симбиотический организм, возникший в результате длительной совместной эволюции гриба и водоросли. Грибной компонент - микобионт - может быть аскомицетом, базидиомицетом или фикомицетом. Фикобионт - сине-зеленые, бурые, золотистые водоросли, но чаще всего зеленые, в том числе почти у половины видов лишайников - зеленая водоросль Тrebouxia. Лишайник отличается особыми способами размножения, медленным ростом, способностью существовать в экстремально бедных местообитаниях, особой биохимией, наличием специфических лишайниковых веществ. Действительно, лишайник можно синтезировать "в культуре", предложив отдельно выращенному микобионту подходящего фикобионта. На том же основан один из способов размножения лишайников, когда выросший из споры микобионт в стадии раннего мицелия должен отыскать рядом подходящего фикобионта и образовать лишайник, если это не удается, гриб погибает. Все остальные способы размножения предусматривают совместное отделение и распространение мико- и фикобионтов. По способу питания все лишайники - автотрофы, никогда не бывают паразитами, "сосущими сок из березы". Ствол березы они могут использовать только как субстрат. "Березовый сок" не соленый, а слегка сладковатый. Это флоэмная жидкость, содержащая мало солей и много сахаров и некоторых других органических веществ. Опята не растут "на травке". Они - древоразрушающие грибы, ксилотрофы. Грибница опят произрастает в ослабленном дереве и постепенно разлагает древесину, питаясь за счет продуктов разложения. Течение "сока" у берез бывает ранней весной, с началом вегетационного периода, а плодовые тела опят появляются поздней осенью, на 5-10 дней, и отмирают. Последняя грубая ошибка в рукописи - цветущий плаун. Плаун - споровое растение, не цветет. "Скромный цвет" - это спорангии, образующиеся на листьях-спорофиллах, которые могут быть собраны в стробилы. Из зрелых спорангиев высыпаются споры, а не семена. Двойственная природа лишайников была открыта в 60-тых годах XIX века немецким ботаником С.Швенденером, а симбиотические отношения в нем были подробно изучены еще позже. С микоризой и микоризообразователями разобрались более-менее лишь к 70-тым годам XX века, хотя как раз об этом автор "рукописи" и не знает. Так что для отставного поручика конца XIX века подделка была совсем, можно сказать, свеженькой.
ВОПРОС 1. "Преждевременная смерть" самцов может быть нужна для того, чтобы они не конкурировали с самками. Например, самцы могут размножаться один раз и погибать, а самки - много. При этом в период между оплодотворением самки и началом развития самцов их (самцов) в популяции нет и плотность её ниже, что снижает конкуренцию за ресурсы. Самки могут размножаться и всего один раз, но после оплодотворения они вынашивают яйца или как-то иначе проявляют заботу о потомстве, после чего умирают; а самцы умирают сразу после оплодотворения. В этом случае конкуренция снижается на время, когда самки заботятся о потомстве. Крайним случаем этого можно считать тот, когда самцы достигают половозрелости гораздо раньше самок, оплодотворяют самок предыдущего поколения и умирают.
Вторым соображением, объясняющим меньшую продолжительность жизни самцов по сравнению с самками, может быть то, что в ситуации, когда самцы размножаются меньшее количество раз, чем самки (не обязательно один, но существенно меньше), быстрее происходит "смена поколений" самцов. Может быть так, что умирая после оплодотворения самец может быть съеден самкой; в таких случаях действует специальный механизм, обеспечивающий то, чтобы во-первых, самца съела оплодотворённая самка, а во-вторых, чтобы она была оплодотворена именно им. Так происходит, например, у пауков.
Наконец, меньший срок жизни самцов по сравнению с самками может и не быть нужен сам по себе, но может являться результатом других действий, имеющих другие цели. Например, если самцы участвуют в брачных турнирах, где могут быть убиты и серьёзно поранены, то средняя продолжительность жизни самцов будет меньше таковой для самок. Это может быть нужно для обеспечения полигинии (ситуации, когда при размножении на одного самца приходится несколько самок). Кроме того, самцы могут вести более открытый образ жизни и из-за этого больше "страдать" от хищников. Это уменьшает пресс хищника на самок и молодняк (см. ответ на вопрос N1 в 9-м классе).
ВОПРОС 2. Причины этого явления могут быть, на наш взгляд, следующими:
1. В норме гусеницы старших возрастов подавляют рост гусениц младших возрастов. Наездник заражает гусениц старших возрастов, они перестают мешать расти гусеницам младших возрастов, чей рост и вызывает большие повреждения капусты.
2. У капустной белянки был вид-конкурент, вредоносное действие которого на капусту было незаметно на фоне белянки. Когда же "благодаря" наезднику численность популяции белянок резко снизилась, увеличилась численность конкурента. К тому же сказалось, что он более эффективно использует капусту как пищевой ресурс, что и приводит к большим её повреждениям.
3. Есть другой наездник (или ещё какой-нибудь паразит), который сильнее влияет на численность капустной белянки, но наездник его легко вытесняет. Вместо "сильного" паразита появляется более "слабый", что вызывает увеличение численности капустной белянки.
4. То, что гусеницы белянки заражены личинками наездника, снижает их пищевую ценность (привлекательность, вкусность) для хищников, например, птиц. Таким образом уменьшается пресс хищников на популяцию белянки, что вызывает увеличение количества гусениц и увеличение повреждений (при этом считается, что зараженная наездником и незараженная им гусеницы поедают капусту с одинаковой или близкой интенсивностью).
5.Каждый год в данной популяции капустной белянки бывала эпизоотия, резко снижавшая численность гусениц. Для того, чтобы эпизоотия "получилась", нужна достаточно большая плотность популяции белянки. Наездник уменьшил её до критического уровня, при котором эпизоотия уже не возникает. Поскольку эпизоотия снимала численновть бабочек больше, чем наездник, получился "выбор из двух зол меньшего".
6. Зараженные наездником гусеницы могут быть более прожорливы, чем незараженные. Пока они не умерли, они будут есть капусту сильнее, чем если бы наездника не было.
7. Птицы, которые обычно питались гусеницами, перешли на питание наездниками и уничтожили большую их часть. В то же время гусеницы выжили и их оказалось больше, чем до применения наездника.
8. Вместо гусениц белянки, которых стало мало, появился другой вредитель, который раньше не питался капустой, и повредил капусту еще больше, чем бы это сделали гусеницы белянки.
Большинство гипотез, приведённых здесь, имеет варианты. Несомненно, есть и другие разумные и непротиворечивые гипотезы, которые могут быть высказаны школьниками и должны быть оценены.
ВОПРОС 3. Функции пигментов бывают самые разные:
1. Пигменты придают окраску шерсти, перьям, чешуе, коже и т. п. животных. О том, для чего бывает нужно, чтобы всё тело или часть его имела какой-нибудь цвет, можно говорить очень долго, но это, увы, не относится к ответу на данный вопрос. Также пигменты придают окраску лепесткам цветов, плодам и т. п. Для придания окраски своему телу можно использовать и чужие пигменты (например, гусеница, съевшая лист какого-то растения, становится зелёной от просвечивающего сквозь кожу и стенки кишечника хлорофилла.
2. Пигменты могут делать непроницаемыми для видимого света отдельные части тела или органы животного, или всё его тело. Например, пигменты радужной оболочки глаза млекопитающих, делая её непрозрачной, позволяют регулировать размер зрачка.
3. Пигменты могут защищать организм, например, от действия ультрафиолетового излучения (они его поглощают). Например, меланин в коже человека защищает от ультрафиолетового излучения глубже лежащие ткани. Иногда такие пигменты локализуются не только в коже, но и в оболочках, окружающих внутреннюю полость организма (например, у пустынных рептилий).
4. Пигменты могут служить для поглощения света, это бывает нужно, например, для фотосинтеза (свет поглощает хлорофилл) или использоваться в зрительной системе (свет поглощается родопсином). Световая энергия в этих и подобных случаях преобразуется в другие виды энергии, которые могут запасаться или использоваться для каких-то специальных целей.
5. Разные вещества, выполняющие в организме разнообразные функции, не связанные с поглощением или отражением света, могут быть окрашенными, то есть являться пигментами. В качестве примеров таких веществ можно привести так называемые дыхательные пигменты, например, гемоглобин, гемоцианин и др. Таким образом, пигменты могут выполнять почти любые функции (то есть все те для которых окраска не является помехой).
ВОПРОС 4. Предпосылкой для всей работы по реконструкции является отнесение данной части по внешним признакам к определенному классу скелетных образований ("зуб", "позвонок" или "раковина") и в тесной взаимосвязи с этим - к определенной, пусть довольно большой систематической группе организмов, вымершей или рецентной. Это дает возможность применить два основных принципа, введенных еще Ж. Кювье: принцип типологической модели и принцип корреляции частей в организме.
В качестве типологической модели может быть использован как ныне живущий, так и вымерший, но уже реконструированный или известный в лучшей сохранности организм, наиболее сходный с реконструируемым по морфологии соответствующих частей. Последнее подразумевает установление гомологий, которое осмысленно лишь в рамках определенной систематической группы и тем самым зависит от предпосылки /от ее верности/. К тому же гомологизация окаменелости может проводиться, как правило, по ограниченному числу признаков /остальные не известны/, а разная изученность разных организмов накладывает ограничения на обоснованный выбор модели. Говоря о корреляции органов, имеют в виду часто их функциональную зависимость, морфофункциональный анализ. Однако далеко не во всех случаях повторяющемуся сочетанию частей - корреляции - мы можем придать определенный функциональный смысл, да и сама функциональная связь может заключаться не в том, что части функционируют совместно, а в том, что они имеют общую морфогенетическую детерминацию /например, могут быть связаны с плейотропным действием генов/. Для реконструкции важно одно - чтобы по присутствию одной части можно было судить или о недостающей части, или о гомологии присутствующей непонятной части. /Отсюда ясно, что первые два принципа должны использоваться совместно - модель подыскивается в расчете на то, что ее части скоррелированы так же, как у реконструируемой формы - и корректируют друг друга и предпосылку, так что весь процесс идет итеративно - путем последовательных приближении/.
Поскольку корреляции отражают не только причинные связи, но и эмпирически установленные соотношения /рогатые млекопитающие непременно должны быть копытными/, да и в функциональных отношениях возможны не всегда известные нам двойственности или непривычные соотношения, здесь возможны ошибки, связанные с наличием в данной систематической группе уклонявшихся по строении и/или образу жизни представителей, недостаточными знаниями о разнообразии модельной группы, преувеличением степени родства с ней реконструируемой формы и т.п.
Так, своеобразный представитель непарнокопытных халикотерий имел на пальцах не копыта, а когти, хотя остальные признаки несомненно указывали на его принадлежность к этой группе. Предполагается, что когти передних конечностей использовались им для пригибания ветвей, с которых он объдал листву, а наличие когтей на задних связано с единством формообразования в онтогенезе этих структур /хотя когти задних лап были менее развиты/. По этой особенности строения халикотерий не имеет аналогов среди других копытных, в том числе современных, и если бы скелет его конечностей не был найден, вряд ли мы могли бы даже вообразить подобное строение его пальцев /а куда могли бы быть отнесены его когти, найденные отдельно!/.
Для реконструкций можно использовать также совпадение ареалов и положения отдельных частей в геологических слоях. Это принцип общей пространственно-временной локализации частей, частным и более ясным случаем которого является совместное нахождение частей в одном захоронении. Единичное совместное захоронение может ничего и не значить, важна многократная повторяемость в разрезе и на площади. Наблюдаемые случаи совместного захоронения отдельных частей, часто в различных сочетаниях, необходимо сравнивать с обычно более редкими находками соединенных между собой частей в тех же слоях с учетом их сочетаемости и морфологической близости /построение новой типологической модели/. Однако надо учитывать, что общая пространственно-временная локализация свойственна и повторяющимся элементам экосистемы (сообщество копытных определенных биотопов данного геологического времени), а также может быть результатом перезахоронений разновременных остатков (в общих осадконакопительных бассейнах и т.п.). Прижизненная связь частей может быть кажущейся, а может быть результатом совместного захоронения разных организмов, связанных биотическими отношениями /например, хищника и жертвы в желудке хищника, или даже отдельных частей жертв в его логове и т.п./.
Знание строения скелета дает представление о пропорциях и общих контурах тела ископаемых животных, а также о строении имевшихся в скелете хрящевых образований, о мускулатуре и других мягких тканях (борозды и отверстия для нервов и кровеносных сосудов, внутренние отливки мозговых капсул черепа, степень развития отдельных мышц определяется размерами, формой и строению поверхности впадин, островков, отростков, бугров и гребней на костях, к которым они крепятся и т.п.). Здесь используются в основном первые два принципа - типологической модели и корреляций, а также общие биомеханические и гистологические соображения. Однако и здесь возможны ошибки, связанные с гипертрофированным развитием отдельных нескелетных образований (хрящей ушных раковин у ряда грызунов или хоботообразных образований непарнокопытных и слонов, игл различной длины и т.п.). Ясно, что дикобраз без игл - уже не дикобраз, а кашалот без огромного спермацетового мешка в передней части головы, размеры которого трудно представимы на основе строения относительно небольших отростков костей, его поддерживающих, был бы на рекунструкции похож на "лобастого" дельфина.
Изучение строения вымерших животных позволяет на основе знания функциональных корреляций судить о наземном, водном или амфибиотическом образе жизни животного, способах его передвижения (вертикальная или горизонтальная постановка хвостового плавника у водных, стопо-, пальце- или фалангохождение у наземных и т.п.), возможностях принимать те или иные цозы /относительная подвижность частей скелета/, пищевой специализации напр., различное строение зубов у хищных и травоядных млекопитающих, а в пределах копытных - у листо- и собственно травоядных форм/ и других особенностях, отражающихся на внешнем облике, поведении и экологии животного. Особенно это важно при последней фазе реконструкции - реконструкции кожи и волосяного покрова, от которых очень редко остаются ископаемые остатки или отпечатки достаточной сохранности и ясности.
Результатом реконструкции является описание, но часто - и изображение внешнего облика и строения животного, поэтому во всех художественных изображениях внешнего облика ископаемых форм присутствует, помимо чисто научного, художественный аспект. В отсутствие однозначных научных данных худэжник-анималист вынужден в определенной степени на свою интуицию полагаться, что часто ведет к увеличению наглядности реконструкции, но уменьшению ее достоверности.
ВОПРОС 5.
а. Разные млекопитающие имеют разные размеры, и потому необходимо разное время, чтобы "построить" их из клеток, имеющих у всех видов примерно одинаковые размеры. С другой стороны, чем крупнее животное, тем дольше оно живет и может позволить себе более длительный срок эмбрионального развития /абсолютно, а не относительно/, чем более мелкие и короткоживущие.
б. У одних видов детеныши рождаются более "доношенные", и они с рождения проявляют большую или меньшую самостоятельность /копытные, зайцы/, у других - менее "доношенные", голые, слепые и беспомощные /напр., крысы/. В первом случае при сходных размерах развитие занимает больше времени, биологический смысл тут такой же, как у птенцовых и выводковых птиц.
в. У сумчатых сама беременность довольно короткая, и рождается еще по существу эмбрион, который доразвивается в сумке.
г. у примитивных млекопитающих /насекомоядные/ беременность длится несколько дольше, поскольку метаболизм происходит медленнее и непостоянна температура тела.
д. У ряда зимоспящих видов развитие замедляется на период спячки и беременность длится дольше.
е. У ряда хищных два сезона гона - осенью и весной, а рожают они только в конце весны, когда достаточна кормообеспеченность. У "осенних" самок беременность тормозится, пока не наступит световой день нужной длины.
ж. У ряда куньих оплодотворяются еще не вполне половозрелые самки, а развитие эмбрионов задерживается до достижения ими размеров взрослых. В крайних случаях, напр., у соболя, самец, найдя гнездо со щенками, оплодотворяет почти новорожденных самок, что избавляет его от необходимости искать их, когда они покинут гнезду и разбегутся.
ВОПРОС 6. Высокая доля вечнозелёных растений в тропиках обеспечивается в первую очередь за счёт влажных дождевых лесов, произрастающих при практически неизменной температуре и влажности. Листопадные растения встречаются и здесь, но зачастую и они стоят зелёными весь год, а листопад происходит у них всё время, но постепенно, то есть не опадают сразу все листья, а выпадают постепенно и по мере опадания заменяются новыми.
В тропических саваннах с их сезонными засухами вечнозелёные растения теряли бы в неблагоприятные периоды слишком много влаги, и поэтому здесь картина иная. Но во-первых, вклад "саванновых" видов растений в общее разнообразие тропических растений мал по сравнению с вкладом растений вечнозелёных лесов, а во-вторых, вечнозелёные" виды есть и в саваннах (кактусы и др. суккуленты, решившие проблему сохранения влаги без листопада /плотные покровы, восковой налёт, преобразование листьев в колючки, запасание воды в стебле и т. д./). В основном это характерно для обитателей мест с очень продолжительным сухим сезоном, когда необходимо испарение снизить, сохранив способность к фотосинтезу. Если сухой сезон длится долго, но осадки в течение него всё-таки выпадают, возникают вечнозелёные жестколистные сообщества вроде субтропических скрэбов.
Резкое уменьшение доли вечнозелёных видов в средних широтах с резкой сменой сезонов и зимними холодами вполне естественно: главные опасности, угрожающие растениям зимой - это высохнуть (если листву не сбрасывать, она будет испарять, а получать воду из почвы растение не может, ибо она там в кристаллическом виде) и для крупных растений - сломаться под тяжестью снега, лежащего на листьях. Поэтов преимущество растений, вегетирующих летом, а на зиму теряющих листву или вообще все надземные органы и зимующих в виде корневищ, клубней и т. п. в умеренных широтах очевидно.
При продвижвнии в арктические широты лето становится всё более коротким и прохладным и в конце концов его начинает не хватать для полноценного наращивания фитомассы, цветения и плодоношения и преимущество опять получают вечнозелёные виды, у которых фитомасса сохраняется несколько лет, а не наращивается каждый год (здесь возникает любопытная аналогия с тропиками, где в районах с засушливым сезоном доля вечнозелёных падает, а в районах с долгим засушливым сезоном - растет). В Арктике играет роль также и то, что лето слишком коротко для однолетников, и они выпадают, что, естественно, повышает долю всех оставшихся в разнообразии и резко уменьшает долю невечнозелёных видов.
Это один взгляд на проблему. На самом деле доля вечнозелёных растений зависит ещё и от того, представители каких групп преобладают в биотопах данной широты. Например, в бореальных районах больше хвойных, чем в средних широтах, а многие хвойные - вечнозелёные.
ВОПРОС 7. Вообще говоря, такая ситуация может сложиться случайно (и это один из вариантов ответа на этот вопрос), но это маловероятно, так как если признак не подвержен отбору, то вряд ли два и более вида будут похожи по этому признаку. Гораздо более вероятны следующие варианты:
1. Несъедобный вид, которому "подражали" съедобные, был, но его не стало. Это могло бы произойти двумя способами: несъедобный вид "выпал" из биоценоза (вымер, ушел и т. д.); появился хищник, для которого несъедобный для других хищников вид съедобен (тогда все рассматриваемые виды съедобны "с точки зрения" этого хищника), при этом "старые" хищники могут как выпадать, так и не выпадать из биоценоза.
2. Эти виды просто родственные. Тогда сходство их внешнего вида совершенно естественно и не требует дополнительных объяснений.
3. Один вид может питаться другим и с этой целью ему подражать. При этом жертва слишком поздно "понимает", что перед ней хищник, а не представитель её вида...
4. Животные разных видов могут "подражать" какому-нибудь неживому объекту, например, листу, веточке и т. п. Поскольку ареалы рассматриваемых видов хотя бы частично перекрываются и среда обитания общая, неудивительно, что "образец для подражания" один.
5. Самки одного вида могут, пользуясь своим сходством с самками другого вида, проникать в гнездо (обычно колониальное) и откладывать туда яйца, снимая тем самым с себя заботу о потомстве и обеспечивая себе большее количество потомков; им также не нужно строить своё гнездо. Так бывает, например, у ос.
6. Колониальные насекомые одного вида, например, муравьи, одного вида могут, пользуясь своим внешним сходством с муравьями другого вида и совершая определённые движения, выпрашивать пищу у особей другого вида.
7. Если из двух похожих видов один редок, а другой массов и они совершают
миграции, собираясь в стаи (так делают некоторые бабочки), то особи редкого
вида могут мигрировать в стаях массового. Это может быть полезно с двух точек
зрения:
1) мигрировать в стае вообще почему-либо лучше, а редкий вид столь редок,
что не может образовывать собственные стаи. Сходство с массовым видом нужно
для того, чтобы в стаю приняли (если есть специальный механизм формирования
стаи, в котором это важно).
2) мигрировать, забираясь в центр стаи (а на периферии - особи массового
вида). Такая организация стаи уменьшает, например, поедание хищниками особей
редкого вида.
Вероятно, при ответе на этот вопрос школьники будут пытаться "обойти" формулировку, например, писать о том, что особи этих видов могут походить на несъедобный вид запахом. Такие "ответы" ответами не считаются.
ВОПРОС 1. Несколько предварительных замечаний:
1. Плотность популяции - число особей на единицу площади. В отсутствие ограничивающих факторов численность популяции увеличивается очень быстро /по экспоненте/, поэтому обсуждаются обычно факторы, ограничивающие увеличение плотности сверху.
2. Плотность определяется соотношением рождаемости, смертности и
миграционного баланса /разницей между вселением (иммиграцией) и выселением
(эмиграцией)/ в популяции. Каждый из этих показателей может зависеть от
плотности (т.е. закономерно изменяться при ее изменении) или не зависеть
(т.е. изменяться случайным по отношению к плотности образом). Последнее
утверждение дает две возможности. Возможно, что каждая популяция обладает
равновесным уровнем плотности и существуют выработавшиеся в ходе эволюции
внутрипопуляционные или биоценотические механизмы, направленные на поддержание
этого уровня (регуляционизм). Тогда при увеличении плотности должна падать
рождаемость, либо увеличиваться смертность, либо миграционный баланс должен
сдвигаться в сторону эмиграции, либо все вместе. Это может происходить через
посредство самых разных явлений: усиления внутривидовой конкуренции /за пищу,
убежища и т.п./, влияния хищников, пресса паразитов, эпизоотий и т.п.
У каждого конкретного вида может быть несколько разных механизмов,
действующих на разных уровнях повышения плотности /см. рисунок/. У животных
с малым числом потомков, длительным репродуктивным циклом и выраженной
заботой о потомстве (К-стратегов) основной механизм - повышение смертности,
особенно среди молодых, при сохранении почти такого же уровня рождаемости
(гибель слетков у синиц при перенаселенности). У животных с большим числом
потомков, коротким репродуктивным циклом и отсутствием заботы о потомстве
(r-стратегов) наряду с повышением смертности наблюдается сильная, иногда
катастрофическая эмиграция /лемминги/ и понижение рождаемости /через меньшую
плодовитость более мелких и менее упитанных в связи с повышением плотности
родителей - у многих насекомых/. Частный случай регуляционной модели -
саморегуляция популяций, когда изменяется не только и не столько качество
среды, сколько качество самих составляющих ее особей. Здесь возможны три
основных механизма торможения роста плотности:
Схема соотношения диапазонов плотности, в пределах которых возможна
регуляция разными биотическими взаимодействиями у животных
(по Г.А.Викторову, 1967 )
1) при возрастании плотности возрастает частота контактов между особями, что приводит к возникновению постоянного стрессового состояния и далее через резкое повышение активности надпочечников при стрессе - к изменению гормонального баланса. Это ведет к повышению смертности через дисфункцию систем органов, а также к снижению рождаемости ввиду более позднего полового созревания, снижения числа зародышей и даже рассасывания эмбрионов /многие грызуны/
2) при возрастании плотности повышением частоты контактов между особями и возрастанием концентрации феромонов усиливается миграция особей из основного местообитания в краевые, где смертность в силу самых разных причин выше /насекомые - саранчовые/; у ряда ящериц к этому же ведет прямая агрессия, связанная с территориальностью и охраной территории
3) при возрастании плотности происходят изменения генетического состава популяции, в частности замена быстро размножающихся генотипов медленно размножающимися и склонными к миграции /полевки, опять же саранчовые и др./. Другая возможность заключается в том, что равновесного уровня не существует (он кажущийся - результат усреднения данных по плотности за длительный срок), как нет и зависимых от плотности факторов его поддержания. Ограничение роста плотности сверху достигается действием случайно распределенных во времени и пространстве неблагоприятных факторов, в основном абиотических (климат), зависящих не от плотности, а от других, обычно глобальных (солнечная активность и др.) причин. В дополнение часто приводится ограниченность ряда ресурсов (пищи, убежищ) также при условии независимого от плотности их использования и распределения. Действительно, такая концепция хорошо описывает динамики численности некоторых животных /особенно массовых видов насекомых, обитающих в изменчивых условиях среды неустойчивых и нарушенных экосистемах/, однако в последнее время она используется большинством экологов только в сочетании с теми или иными вариантами первой концепции (комплексность механизмов регуляции).
Обе концепции сходятся в том, что наблюдаемые постоянные колебания численности и в меньшей степени - плотности есть результат запаздывания реакции ограничивающих факторов на изменение численности и плотности.
ВОПРОС 2. Защита бактерий от вирусов предусматривает специальные приспособления, с помощью которых бактерия эффективно защищается от вирусов. Вариант ответов типа "бактерия подавляет синтез собственных нуклеиновых кислот и белка, не давая вирусам размножаться" не удовлетворительны, поскольку в результате такой защиты клетка гибнет (либо должны приводиться убедительные причины того, почему она все же не гибнет, или почему гибель отдельных /именно отдельных!/ клеток может быть защитой для популяции в целом). Не годятся также ответы типа "бактерии стараются жить там, где никаких вирусов нет" или версии, предусматривающие приспособления, которых нет у бактерий, например ядерную оболочку или зрительную систему, которая позволяет бактерии вовремя обнаружить вирус и дает счастливую возможность убежать от него при помощи своих или чужих ног.
Остальные версии делятся на две большие группы: защита от проникновения вируса и защита от его размножения и лизиса клетки тогда, когда он уже проник. Основные идеи:
1) Проникновение бактериального вируса (бактериофага, или просто фага) в бактерию начинается с того, что он связывается с определенным белком-рецептором, находящимся в мембране бактериальной клетки или в ее клеточной стенке. Устойчивость к заражению фагами некоторых штаммов бактерией обусловлено, вероятно, отсутвием у них рецепторов для данных фагов. Далее ряд вирусов растворяет с помощью одного из собственных белков клеточную стенку и при взаимодействии с рецептором и мембраной образует канал, через который нуклеиновая кислота фага проникает внутрь клетки. Значит, устойчивость бактерий к разным фагам может определяться особенностями химического строения клеточной стенки. Другие вирусы, по всей вероятности, не могут растворять клеточную стенку, и проникают в бактерию через лишенные оной малые по площади мембранные выросты - пили (используемые бактерией, в частности, для конъюгации). Объединенные в колонии бактерии экранируют от вируса рецепторы и пили, а если крайние клетки поражаются вирусом и лизируются, обрывки их оболочек продолжают отчасти экранировать оставшиеся клетки. Два способа защиты на расстоянии: выделение слизистых и иных защитных капсул и выделение в первую очередь для внешнего пищеварения расщепляющих белки ферментов, которые попутно могут расщеплять и белки оболочки вирусов.
2) Внутри бактериальной клетки существуют ферменты рестриктазы, умеющие отличать хозяйские нуклеиновые кислоты (некоторые нуклеотиды которых химически изменены специфическим для данного штамма образом) от любых чужеродных (происходящих не из клеток данного штамма) и расщепляющие последние. Однако если вирус уже размножался в клетках данного штамма, его ДНК или РНК имеет такое же строение.
3) Существуют так называемые умеренные бактериофаги, ДНК которых встраивается в геном бактерии и длительное время не экспрессируется, вследствие чего лизиса не происходит, и фаг (называемый в этом случае профагом) размножается только синхронно с размножением бактериальной клетки. Это состояние обусловлено действием белка-репрессора, подавляющего транскрипцию ДНК профага. Поэтому клетки, зараженные умеренным фагом в форме профага, устойчивы к любому последующему заражению этим фагом. При некоторых воздействиях и внутренных процессах репрессор, однако, инактивируется, вызывает тем самым размножение фага и лизис клетки. Ряд штаммов бактерий имеют в геноме "сломанный" профаг, не способный активироваться, и обладают высокой устойчивостью к заражению этим фагом.
ВОПРОС 3. Возможно, что обратная транскриптаза /далее - ОТ/ принадлежит не собственно эукариотической клетке, а особой группе поражающих их вирусов (ретровирусам). Эти вирусы в качестве информационной молекулы содержат одноцепочечную РНК, один из генов в которой и кодирует ОТ. При попадании вируса в клетку сначала с вирусной РНК транслируется ОТ, затем ОТ на матрице вирусной РНК синтезирует двухцепочечную ДНК, одна из цепей которой соответсствует, а другая - комплементарна матрице, и далее специальные ферменты в клетке /сайт-специфические нуклеазы/ встраивают эту ДНК между определенными участками клеточной ДНК /сайтами/, или, что бывает чаще, другие клеточные нуклеазы втраивают эту ДНК в разные места клеточного генома случайным образом. Далее в большинстве мест вирус будет "молчать", его гены не будут экспрессироваться, и клетка проживет положенный срок, передавая встроенный вирус своим потомкам /а если клетка половая - то и целым организмам/.
Лишь встроившись в определенное или даже единственное место в геноме, либо попав в определенную ткань /в то время как имеется в неактивной форме и в других тканях/ вирус обретает способность размножиться и выйти из клетки. В первом случае это определяется особенностями соседних с вирусом участков клеточной ДНК, а во втором - внешними специфическими для этой ткани воздействиями /например ретровирус молочной железы мышей содержится во всех тканях больного животного, но усиленно транскрибируется и образует вирусные частицы только в ткани молочной железы у мышей определенного возраста. Это связано с наличием именно в этой ткани /в ее клетках/ специфических белков рецепторов для ряда половых гормонов, вызывающих созревание клеток железы у мышей определенного возраста. Гормоны свободно проникают через липидную мембрану клеток, связываются в цитоплазме с белками-рецепторами и в составе этого комплекса присоединяются к ДНК клетки для регуляции транскрипции некоторых клеточных генов, продукты которых и вызывают созревание клеток железы. Это, таким образом, нормальный процесс. Но поскольку место их присоединения оказывается очень близко к промотору (регуляторной последовательности) вирусной ДНК, это одновременно активирует промотор и начинается экспрессия вирусного генома и размножение вируса /синтез вирусных РНК и белков, в том числе ОТ/.
Бывает, что ретровирусы теряют способность нормально размножаться и выходить из клетки вследствие мутаций одного или нескольких белков их оболочки /не происходит самосборки/, но другие их гены, и в том числе ген ОТ, могут нормально транскрибироваться и транслироваться при указанных выше условиях. Возможно, что именно в этом причина того, что из клеток все большего числа эукариот выделяется обратная транскриптаза при отсутствии признаков вирусной инфекции. Вероятно также, что она там каким-то образом функционирует, поскольку встречается в цитоплазме в комплексе с клеточной РНК и белками.
В чем смысл ее активности - пока достоверно не известно, что, однако, никак не мешает школьникам выдвигать более или менее обоснованные гипотезы на основе знания ее функции как фермента и основных закономерностей устроцйства и работы генома и синтетического аппарата клетки. Возможны такие основные группы идей:
1) группа ответов, связанных с амплификацией (увеличением числа копий)
генов в геноме. ОТ синтезирует копии гена на матрице и-РНК этого гена,
и далее эти копии встраиваются в геном. Это может приводить:
а. К каскадному увеличению числа копий гена, что может быть важно при
необходимости синтеза большого количества молекул белка /напр., при секреции/
или РНК /рибосомальных и транспортных/. Реально в геноме эукариот
действительно встречаются высокоповторяющиеся последовательности (от ста
тысяч до миллиона копий на геном), умеренно повторяющиеся (от ста до десяти
тысяч копий на геном) и уникальные (до 10 копий на геном).
б. копии могут встраиваться в тех же местах, что и ранее существующие гены, а могут и в другие места генома, в том числе в разные хромосомы, что
может приводить к увеличению рекомбинации или изменению групп сцепления.
Такие события должны быть достаточно редкими - иначе не выполнялись бы
законы наследования.
в. В геноме эукариот есть транскрибируемые и нетранскрибируемые участки,
а в первых - кодирующие и некодирующие /гл. обр. регуляторные/ участки
(экзоны и интроны). Последние вырезаются ферментами из первичного транскрипта
в ходе его созревания - процессинга, происходящего в ядре, и в цитоплазму
поступают не содержащие этих участков и-РНК. Поскольку в ходе процессинга
вырезается до 55% длины первичного транскрипта, можно предположить, что
наличие в геноме синтезированных ОТ копий без интронов позволило бы
увеличить скорость синтеза генных продуктов, обойдясь без процессинга
/который занимает, конечно, значительное время/, а заодно навести экономию
генного материала. Реально это натыкается на трудно преодолимые препятствия:
как тогда обеспечивать регуляцию транскрипции и как обеспечить стабильность
и-РНК (устойчивость к расщеплению нуклеазами, достигаемая в норме
добавлением с обоих концов транскрипта определенных нуклеотидов и
модификацией, напр., метилированием, некоторых оснований внутри транскрипта -
все это достигается в ходе процессинга).
2) группа ответов, связанных с дополнительными механизмами мутаций (общая идея в том, что изменения в РНК после транскрипции не утрачиваются, а от ражаются в геноме в результате активности ОТ)
3) группа ответов, связанная с отбором более стабильных (долгоживущих) РНК, так как с них может быть сделано больше копий
4) группа ответов, связанных с использованием возникающего в ходе работы ОТ комплекса РНК-ДНК для восстановления нарушений в ДНК генома, и т.п.
В целом очевидно, что данный вопрос является по сути лишь затравкой для выяснения глубины и правильности понимания школьниками процессов синтеза нуклеиновых кислот и регуляции генной экспрессии, и его проверка должна осуществляться именно с этих позиций.
ВОПРОС 4. Все гипотезы о функциях генов А, В и С можно расположить в рамках двух моделей: мультисигнальной и унисигнальной. Вообще говоря, можно придумать гораздо больше способов формирования цветка, удовлетворяющих условию задачи, чем приводим мы. Данный вопрос тем и интересен, что проверяет способности школьников строить разнообразные, более или менее глубокие модели исходя из определенных условий.
Итак, мультисигнальная модель. Она подразумевает, что формирование разных органов цветка обусловлено действием на меристему разных сигналов (веществ, способных вызывать дифференцировку клеток по определенному типу).
Гипотеза N1. Сигналы выделяются какими-то клетками на апикальном конце меристемы /формирование цветка идёт "сверху вниз", т. е., первыми образуются плодолистики, а последней - чашечка/. Гены А, В и С ответственны за синтез (или выделение, или синтез предшественников сигналов и вообще за любой процесс, результатом которого является выделение сигнала). В дальнейшем для краткости будем писать, что гены ответственны за результат, подразумевая при этом, что их непосредственная функция может быть другой. В разное время (последовательность выделения их указывают цифры возле вертикальной оси.
Результаты мутаций в парах генов:
А и В - 1 круг - чашелистики (чшл)
2 круг - чшл
3 круг - чшл
4 круг - плодолистики (пдл)
А и С - 1 круг - чшл
2 круг - чшл
3 круг - тычинки (тч)
4 круг - тч
В и С - 1 круг - чшл
2 круг - лепестки (лп)
3 круг - тч
Результат мутаций во всех трёх генах:
1 круг - чшл,
2 круг - чшл,
3 круг - чшл
4 круг - чшл
Гипотеза N2. Выделяется 3 сигнала, одинаковых по силе, в разное время. Гены определяют восприимчивость клеток определенных групп к сигналу.
Последовательность выделения сигналов
указывают цифры возле вертикальной оси.
Результаты мутаций в парах генов и мутаций
во всех трех генах предлагается предсказать.
При этом будут появляться результаты типа
1 круг - чшл, 2 круг - чшл, 3 круг - чшл, 4 круг - лп.
Противоречие условию задачи здесь можно устранить путем следующих рассуждений: "мутация в гене А вызывает развитие чшл вместо лп не только в "своём" круге но и в любом другом круге цветка. Так же и с мутациями в генах В и С."
Гипотеза N3. Сигналы выделяются в основании меристемы, а апикальный её
кончик выделяет ингибиторы, сводящие действие сигналов на нет (при этом
формирование цветка идёт, конечно, "снизу вверх", т. е. первыми закладываются
чшл, а пдл - последними. В таком порядке происходит формирование органов
цветка на самом деле.). Сигналы одинаковой силы вырабатываются в разное
время; граница зоны, в которой они работают, определяется границей зоны
проникновения ингибитора, которая зависит от его количества, а это
количество регулируется генами А, В и С,
Теперь перейдём к унисигнальной модели. Здесь мы ограничимся одной
гипотезой. Основание меристемы выделяет некое вещество, концентрация
которого линейно убывает по мере удаления от основания меристемы. Результат
клеточной дифференцировки зависит от концентрации вещества, выделяемого
основанием меристемы (сигнала) следующим образом: при концентрации в-ва
(силе сигнала) ниже определенного уровня формируются чшл, от одного уровня
до другого - лп и т. д. Гены А, В, С определяют эти "критические уровни".
При мутации в каком-то из этих генов соответствующий порог ("критический
уровень" перестает существовать и продолжается нормирование элементов
предыдущего круга до тех пор, пока не вступит в действие продукт ближайшего
"по порядку" нормального гена. Результаты мутаций в парах генов также
предлагается предсказать.
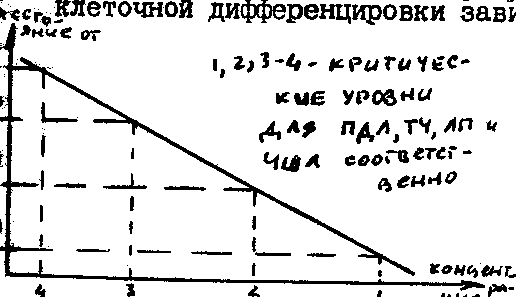
ВОПРОС 5. Размер одноклеточного организма ограничивают сверху следующие факторы:
А. Возможность обмена с окружающей средой, т.к. при увеличении линейных размеров объем возрастает пропорционально кубу, а поверхность - лишь квадрату линейных размеров.
Б. Так как скорость диффузии молекул не зависит от размеров клетки, то у крупных клеток возникают сложности с доставкой необходимых веществ в нужную точку, т.е. с транспортом.
В. Ядро может "обслужить" своими продуктами ограниченную часть цитоплазмы, и ядерно-цитоплазматическое соотношение довольно постоянно в больших таксонах организмов.
Г. Сложность поддержания формы - чем больше клетка, тем больше механические напряжения в ней и слабее действие сил поверхностного натяжения окружающей жидкости, которые в значительной степени поддерживают округлокаплевидную форму мелких организмов. Одновременно с увеличением размеров возрастает опасность повреждения внешних покровов, которое чревато гибелью всего организма /а не отдельной части, как у многоклеточных/.
Д. Сложности с движением, поскольку мощность движителей должна увеличиваться в соответствии с увеличением массы /объема/ в кубической степени, а площадь приложения движущих сил /она же площадь размещения многих движителей - напр., жгутиков, она же площадь поверхности/ увеличивается пропорционально квадрату, да еще используется для обмена со средой (заметим в скобках, что увеличение скорости движения улучшает обмен, поскольку преодолевая за единицу времени вдвое большее расстояние организм как бы имеет за ту же единицу времени вдвое большую поверхность). В связи с этим линейные размеры движителей должны увеличиваться диспропорционально увеличению линейных размеров клетки в целом, а это не так то просто, в частности потому, что многие их структурные элементы /микротрубочки, базальные тельца жгутиков и др./ имеют фиксированное строение у всех организмов в соответствии с уникальными механизмами их молекулярной сборки в клетке. С возрастанием массы уменьшается также плавучесть, позволяющая двигаться с токами жидкости /пассивно/. Возрастают также удельные затраты энергии на движение.
Е. Возникают также трудности с передачей сигналов и вообще с координированным действием частей оольшой клетки-организма.
Ж. Более общим путем преодоления указанных затруднений является формирование у одноклеточных колониальности, оптимизирующей положение клеток в пространстве, и многоклеточности, связанной также с дифференцировкой. Из 27 типов простейших по одной из последних классификаций в 16 наряду с "истинно" одноклеточными формами присутствуют также более или менее колониальные или многоклеточные формы, некоторые из которых по уровню интеграции клеток не уступают, например, губкам. Но в вопросе спрашивается именно о крупных одноклеточных, их то мы и рассмотрим. При этом одноклеточным следует считать организм, цитоплазма которого едина, причем если ядер много, то они потенциально способны к миграции по всем частям организма, содержащим ядра. К числу очень крупных форм среди одноклеточных /от 2 мм и более, до нескольких см/ можно отнести хитридиомицетов, оомицетов, часть зигомицетов, большинство миксомицетов (все эти формы обычно относят к грибам); всех миксоспоридий; немногие жгутиконосцы из динофлагеллят /напр., ночесветка/; наиболее крупные грегарины, амебы, радиолярии, солнечники, фораминиферы и инфузории; некоторые сифоновые /ацетабулярия, бриопсис, галицистис/ и вошериевые /единственный род - Vauscheria/ водоросли. Необходимую информацию о них можно почерпнуть из книг: Догель В.А. Зоология беспозвоночных. М.: Высш. школа, 1981 г., Курс низших растений. Под ред. М.В.Горленко. М., Высшая школа, 1981 г.; К.Хаусман. Протозоология: Пер. с нем.- М.,Мир, 1988 г.
Здесь приводятся только основные пути ослабления ограничений на размеры:
А. Сокращение относительной поверхности тела отчасти восполняется тонкими цитоплазматическими выростами и общим уплощением тела клетки /саркодовые, грибы/, высокой скоростью движения /инфузории/ или очень высокой плотностью хлоропластов в пристеночном слое /ботридиум/.
Б. Создание специальных токов цитоплазмы в клетке с затратой энергии АТФ /с помощью цитоскелета/, транспорт таким же способом митохондрий и пластид; другие способы решения проблемы приведены ниже.
В. Разные формы усиления функции ядра: много одинаковых ядер /опалины, амебы, примитивные инфузории, грибы, водоросли/, одно высокополиплоидное ядро /ацетабулярия, радиолярии/, разноядерность у инфузорий, при которой микронуклеусы обеспечивают сохранение и передачу информации, а макронуклеусы состоят из большого /до миллиона/ - числа копий генов, необходимых для обслуживания клетки (они составляют небольшой % от имеющихся в микронуклеусе генов). Распределение ядер по клетке решает отчасти проблемы транспорта и передачи сигналов.
Г. Разные типы внутреннего и внешнего скелетов: аксостили, пелликула, белковые, целлюлозные, хитиновые, кремнеземные, известковые и гликопротеидные панцыри и раковины. Сочетание быстрого движения и наличия пелликулы при гетеротрофности привело к возникновению у инфузорий "рта" /цитостома/. Раковина фораминифер пронизана цитоплазматическими выростами; это, наряду с внешними разветвленными ризоподами, компенсирует уменьшение открытой поверхности тела, позволяя достигать огромных размеров /до 145 мм в диаметре раковины/.
Д. Проблема движения решается по разному: полимеризацией двигательных структур /инфузории, опалиниды/, паразитическим образом жизни /миксоспоридии, грегарины/, отказом от больших горизонтальных перемещений в пользу вертикального путем изменения плавучести /радиолярии, фораминиферы, солнечники, которые увеличивают плавучесть выростами скелета и цитоплазмы, запасанием веществ легче воды, созданием в раковине воздушных камер и пр./, постоянным врастанием в среду и разветвленностью тела /грибы/. Рост растений и очень медленное движение амеб также можно рассматривать как врастание в среду.
Е. Для передачи сигналов используются "быстрые" способы типа поляризации. мембран. Вместе с тем создаются условия для самостоятельного реагирования каждым участком /например, грибы при возникновении повреждений в участке мицелия обычно просто отгораживают поврежденный участок от остального мицелия клеточной стенкой/.
ВОПРОС 6. Основные идеи:
а. изменение распределения пигмента в содержащих их клетках - хроматофорах. Когда пигмент в виде мелких гранул рассеян по цитоплазме, окраска кажется более интенсивной, чем когда он собран в центре клетки. Перераспределение пигмента производится с помощью элементов цитоскелета при воздействии на клетку некоторых химических веществ, света, температуры и пр.
б. хроматофоры могут быть распределены по коже неравномерно. Тогда при образовании складок они могут оказываться либо снаружи, либо внутри, что и обеспечивает изменение окраски.
в. Если хроматофор содержит несколько пигментов, то их гранулы могут распределяться в разных сочетаниях и/или экранировать гранулы другого цвета, как это бывает у палочников
г. Хроматофоры могут вытягивать свои отростки поверх соседних, иначе окрашенных клеток и увелмчивать площадь этих отростков, либо наоборот. Иногда таким образом могут взаимодействовать несколько слоев хроматофоров с разными пигментами. Наряду с а) это составляет основу изменения окраски у кальмаров, донных рыб), древесных лягушек, ящериц и управляется нервной системой и антагонистическими гормонами гипофиза и эпифиза.
д. Светящиеся животные могут менять окраску, регулируя свое свечение разными способами /"зашторивание" светящегося органа, биохимическая регуляция люциферин-люциферазной системы, стрессирование светящихся симбионтов изменением рН в местах их локализации и пр./
е. Прозрачные и полупрозрачные животные могут менять окраску при поедании окрашенной пищи, частичном или полном переваривании придающих им цвет симбионтов и т.п.
ж. Прилив крови к коже под действием гормонов /норадреналин - когда человек краснеет/ или изменения температуры изменяет ее окраску.
з. Один из самых простых способов - поведенчески /вываливание в грязи, намокание с потемнением, самомаскировка у крабов и пр./
и. Второй простой способ - линька. Новые верхние слои кожи могут иметь иной цвет гл. обр. из-за иного распределения пигмента.
Возможны и другие разумные идеи. Все перечисленные выше реально существуют.
ВОПРОС 7. Основные идеи:
1) сцепленное наследование данного признака с геном, локализация которого известна, или с полом;
2) корреляция случаев аномального проявления или аномального наследования этого признака с необычным количеством или необычной морфологией какой-то из хромосом
3) активный синтез РНК на какой-то хромосоме непосредственно перед проявлением данного признака. При использовании специальных способов выделения и окраски хромосом это может быть видно в световой микроскоп.
4) если известна последовательность гена, отвечающего за данный признак (возможно - у родственного вида), то его можно локализовать по связыванию с комплементарной цепью ДНК (или РНК).
5) можно также выделить РНК из рибосом, на которых активно синтезируются белок - продукт данного гена, и выявить хромосому с помощью такого РНК-зонда. Просто по строению белка однозначно определить кодирующую его ДНК нельзя, т.к. одна аминокислота может соответствовать различным триплетам ДНК, а в "подозрительных" участках ДНК не известна рамка считывания.